Hebb則と教師なし学習
Contents
Hebb則と教師なし学習#
Hebb則#
神経回路はどのようにして自己組織化するのだろうか.1940年代にカナダの心理学者Donald O. Hebbにより著書”The Organization of Behavior”[Hebb, 1949] で提案された学習則は「細胞Aが反復的または持続的に細胞Bの発火に関与すると,細胞Aが細胞Bを発火させる効率が向上するような成長過程または代謝変化が一方または両方の細胞に起こる」というものであった.すなわち,発火に時間的相関のある細胞間のシナプス結合を強化するという学習則である.これをHebbの学習則 (Hebbian learning rule) あるいはHebb則(Hebb’s rule) という.Hebb則は (Hebb自身ではなく) Shatzにより”cells that fire together wire together” (共に活動する細胞は共に結合する)と韻を踏みながら短く言い換えられている [Shatz, 1992].
Hebb則の導出#
数式でHebb則を表してみよう.\(n\)個のシナプス前細胞と\(m\)個の後細胞の発火率をそれぞれ\(\mathbf{x}\in \mathbb{R}^n, \mathbf{y}\in \mathbb{R}^m\) とする.前細胞と後細胞間のシナプス結合強度を表す行列を\(\mathbf{W}\in \mathbb{R}^{m\times n}\)とし,\(\mathbf{y}=\mathbf{W}\mathbf{x}\)が成り立つとする.このようなモデルを線形ニューロンモデル (Linear neuron model) という.このとき,Hebb則は
として表される.ただし,\(\tau\)は時定数であり,\(\eta:=1/\tau\) は学習率 (learning rate) と呼ばれる,学習の速さを決定するパラメータとなる.
このHebb則は数学的に導出されたものではないが,特定の目的関数を神経活動及び重みを変化させて最適化するようなネットワークを構築すれば自然に出現する.このようなネットワークをエネルギーベースモデル (energy-based models) といい,次章で扱う.エネルギーベースモデルでは,先に目的関数 \(\mathcal{L}\) を定義し,その目的関数を最小化するような神経活動 \(\mathbf{z}\) および重み行列 \(\mathbf{W}\) のダイナミクスをそれぞれ,
として導出する.この手順の逆を行う,すなわち先に神経細胞の活動ダイナミクスを定義し,神経活動で積分することで神経回路の目的関数\(\mathcal{L}\)を導出でき,さらに\(\mathcal{L}\)を重み行列で微分することでHebb則が導出できる [Isomura and Friston, 2020].Hebb則の導出を連続時間線形ニューロンモデル \(\dfrac{d\mathbf{y}}{dt}=\mathbf{W}\mathbf{x}\) を例にして考えよう.ここで\(\dfrac{\partial\mathcal{L}}{\partial\mathbf{y}}:=-\dfrac{d\mathbf{y}}{dt}\)となるような目的関数 \(\mathcal{L}\in \mathbb{R}\)を仮定すると,
となる.これをさらに\(\mathbf{W}\)で微分すると,
となり,Hebb則が導出できる (簡単のため時定数は1とした).
Hebb則の安定化とLTP/LTD#
BCM則#
Hebb則には問題点があり,シナプス結合強度が際限なく増大するか,0に近づくこととなってしまう.これを数式で確認しておこう.前細胞と後細胞がそれぞれ1つの場合を考える.2細胞間の結合強度を\(w\ (>0)\) とし,\(y=wx\)が成り立つとすると,Hebb則は\(\dfrac{dw}{dt}=\eta yx=\eta x^2w\)となる.この場合,\(\eta x^2>1\) なら \(\lim_{t\to\infty} w= \infty\), \(\eta x^2<1\) なら \(\lim_{t\to\infty} w= 0\) となる.当然,生理的にシナプス結合強度が無限大となることはあり得ないが,不安定なほど大きくなってしまう可能性があることに違いはない.このため,Hebb則を安定化させるための修正が必要とされた.
Cooper, Liberman, Ojaらにより頭文字をとってCLO則 (CLO rule) が提案された [Cooper et al., 1979].その後,Bienenstock, Cooper, Munroらにより提案された学習則は同様に頭文字をとってBCM則 (BCM rule) と呼ばれている[Bienenstock et al., 1982] [Cooper and Bear, 2012].
\(\mathbf{x}\in \mathbb{R}^d, \mathbf{w}\in \mathbb{R}^d, y\in \mathbb{R}\)とし,単一の出力\(y = \mathbf{w}^\top \mathbf{x}=\mathbf{x}^\top \mathbf{w}\)を持つ線形ニューロンを仮定する.重みの更新則は次のようにする.
ここで関数\(\phi\)は\(\phi(y, \theta_m)=y(y-\theta_m)\)などとする.また\(\theta_m:=\mathbb{E}[y^2]\)は閾値を決定するパラメータ,修正閾値(modification threshold) であり,
として更新される.
ToDo: 詳細
using PyPlot, Random, Distributions, LinearAlgebra, FFTW
rc("axes.spines", top=false, right=false)
rc("font", family="Meiryo")
ϕ(y, θₘ) = y * (y - θₘ);
y = 0:0.1:2;
θₘ = 1.0
props = Dict("boxstyle" => "round", "facecolor" => "wheat", "alpha" => 0.5)
figure(figsize=(3, 2))
plot(y, ϕ.(y, θₘ))
xlim(0,)
annotate(text="", xy=(0.8,1), xytext=(1.2,1), arrowprops=Dict("arrowstyle" => "<->", "color" => "tab:purple"))
axvline(θₘ, linestyle="dashed", color="tab:purple")
axhline(0, linestyle="dashed", color="tab:gray")
xticks([]); xlabel(L"$y$ "*"(シナプス後細胞の活動)")
ylabel(L"$\phi(y, \theta_m)$")
text(0.7, 1.2, L"\theta_m", color="tab:purple",fontsize=11)
text(0.5, 1.8, "LTD",fontsize=11, color="tab:blue",ha="center",va="center", bbox=props);
text(1.5, 1.8, "LTP",fontsize=11, color="tab:red",ha="center",va="center", bbox=props);
tight_layout()
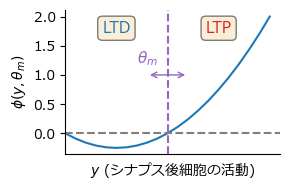
Hebb則の生理的機序#
ここでHebb則およびBCM則の生理的基盤について触れておこう. LTPの実験的発見 [Bliss and Lomo, 1973] [Dudek and Bear, 1992]
ToDo:実験的発見のsurvey
Oja則#
Hebb則を安定化させる別のアプローチとして,結合強度を正規化するという手法が考えられる.BCM則と同様に\(\mathbf{x}\in \mathbb{R}^d, \mathbf{w}\in \mathbb{R}^d, y\in \mathbb{R}\)とし,単一の出力\(y = \mathbf{w}^\top \mathbf{x}=\mathbf{x}^\top \mathbf{w}\)を持つ線形ニューロンを仮定する.\(\eta\)を学習率とすると,\(\mathbf{w}\leftarrow\dfrac{\mathbf{w}+\eta \mathbf{x}y}{\|\mathbf{w}+\eta \mathbf{x}y\|}\)とすれば正規化できる.ここで,\(f(\eta):=\dfrac{\mathbf{w}+\eta \mathbf{x}y}{\|\mathbf{w}+\eta \mathbf{x}y\|}\)とし,\(\eta=0\)においてTaylor展開を行うと,
ここで\(\|\mathbf{w}\|=1\)として,1次近似すれば\(f(\eta)\approx \mathbf{w} + \eta \left(\mathbf{x}y-y^2 \mathbf{w}\right)\)となる.重みの変化が連続的であるとすると,
として重みの更新則が得られる.これをOja則 (Oja’s rule) と呼ぶ [Oja, 1982].こうして得られた学習則において\(\|\mathbf{w}\|\to 1\)となることを確認しよう.
より,\(\dfrac{d\|\mathbf{w}\|^2}{dt}=0\)のとき,\(\|\mathbf{w}\|= 1\)となる.
恒常的可塑性#
Oja則は更新時の即時的な正規化から導出されたものであるが,恒常的可塑性 (synaptic scaling)により安定化しているという説がある[Turrigiano, 2008][Yee et al., 2017].しかし,この過程は遅すぎるため,Hebb則の不安定化を安定化するに至らない[Zenke et al., 2017]
ToDo:恒常的可塑性の詳細
Johansen, Joshua P., Lorenzo Diaz-Mataix, Hiroki Hamanaka, Takaaki Ozawa, Edgar Ycu, Jenny Koivumaa, Ashwani Kumar, et al. 2014. “Hebbian and Neuromodulatory Mechanisms Interact to Trigger Associative Memory Formation.” Proceedings of the National Academy of Sciences 111 (51): E5584–92.
Hebb則と主成分分析#
Oja則を用いることで主成分分析(Principal component analysis; PCA) という処理をニューラルネットワークにおいて実現できる.主成分分析とは-
ToDo:主成分分析の説明
d = MvNormal([0,0], [1.0 0.5; 0.5 1.0]) # multivariate normal distribution
N = 300 # sample size
Random.seed!(0) # set seed
X = rand(d, N); # generate toy data
U, S, V = svd(X*X')
SVD{Float64, Float64, Matrix{Float64}, Vector{Float64}}
U factor:
2×2 Matrix{Float64}:
-0.722509 -0.691362
-0.691362 0.722509
singular values:
2-element Vector{Float64}:
418.9073852600819
138.22321877667497
Vt factor:
2×2 Matrix{Float64}:
-0.722509 -0.691362
-0.691362 0.722509
figure(figsize=(3,3))
scatter(X[1,:], X[2,:], alpha=0.5)
arrow(0, 0, V[1,1], V[2,1], head_width=0.2, color="tab:red", length_includes_head=true, label="PC1")
arrow(0, 0, V[1,2], V[2,2], head_width=0.2, color="tab:orange", length_includes_head=true, label="PC2")
θc = 0:1e-2:2pi
plot(cos.(θc), sin.(θc), "k--", alpha=0.8)
xlabel(L"$X_1$"); ylabel(L"$X_2$")
legend(); tight_layout()
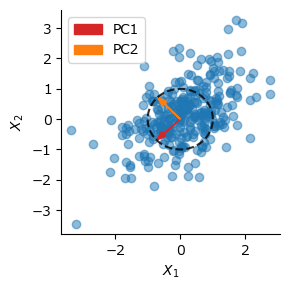
Oja則によるPCAの実行#
ここでOja則が主成分分析を実行できることを示す.重みの変化量の期待値を取る.
\(\mathbf{C}:=\mathbb{E}[\mathbf{x}\mathbf{x}^\top]\in \mathbb{R}^{d\times d}\)とする.\(\mathbf{x}\)の平均が0の場合,\(\mathbf{C}\)は分散共分散行列である.\(\mathbb{E}\left[\dfrac{d\mathbf{w}}{dt}\right]=0\)となる\(\mathbf{w}\)が収束する固定点(fixed point)では次の式が成り立つ.
これは固有値問題であり,\(\lambda:=\mathbf{w}^\top \mathbf{C} \mathbf{w}\)は固有値,\(\mathbf{w}\)は固有ベクトル(eigen vector)になる.
ここでサンプルサイズを\(n\)とし,\(\mathbf{X} \in \mathbb{R}^{d\times n}, \mathbf{y}=\mathbf{X}^\top\mathbf{w} \in \mathbb{R}^n\)とする.標本平均で近似して\(\mathbf{C}\simeq \mathbf{X}\mathbf{X}^\top\)とする.この場合,
となる.
w = randn(2) # initialize weight
w ./= sqrt.(sum(w.^2)) # L2 normalize
initw = copy(w) # save initial weight
η = 1e-3 # learning rate
for _ in 1:200
y = X' * w
w += η * (X * y - y' * y * w) # Oja's rule
end
figure(figsize=(3,3))
scatter(X[1,:], X[2,:], alpha=0.5)
arrow(0,0,initw[1],initw[2], head_width=0.2, color="k", length_includes_head=true, label=L"Init. $w$")
arrow(0,0,w[1],w[2], head_width=0.2, color="tab:red", length_includes_head=true, label=L"Opt. $w$")
plot(cos.(θc), sin.(θc), "k--", alpha=0.8)
xlabel(L"$X_1$"); ylabel(L"$X_2$")
tight_layout()
legend(); tight_layout()
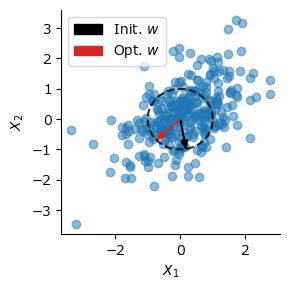
後のためにOja則においてネットワークが\(q\)個の複数出力を持つ場合を考えよう.重み行列を\(\mathbf{W} \in \mathbb{R}^{q\times d}\), 出力を\(\mathbf{y}=\mathbf{W}\mathbf{x} \in \mathbb{R}^{q}, \mathbf{Y}=\mathbf{W}\mathbf{X} \in \mathbb{R}^{q\times n}\)とする.この場合の更新則は
となる.ただし,\(\mathrm{Diag}(\cdot)\)は行列の対角成分からなる対角行列を生み出す作用素である.
Sanger則#
Oja則に複数の出力を持たせた場合であっても,出力が直交しないため,PCAの第1主成分しか求めることができない.Sanger則 (Sanger’s rule),あるいは一般化Hebb則 (generalized Hebbian algorithm; GHA) は,Oja則にGram–Schmidtの正規直交化法(Gram–Schmidt orthonormalization) を組み合わせた学習則であり,次式で表される.
\(\mathrm{LT}(\cdot)\)は行列の対角成分より上側の要素を0にした下三角行列(lower triangular matrix)を作り出す作用素である.Sanger則を用いればPCAの第2主成分以降も求めることができる.
W = randn(2, 2) # initialize weight
W ./= sqrt.(sum(W.^2, dims=2)) # normalize
initW = copy(W) # save initial weight
for _ in 1:200
Y = W * X
W += η * (Y * X' - LowerTriangular(Y * Y') * W) # Sanger's rule
end
figure(figsize=(3,3))
scatter(X[1,:], X[2,:], alpha=0.5)
arrow(0, 0, W[1,1], W[1,2], head_width=0.2, color="tab:red", length_includes_head=true, label=L"$w_1$")
arrow(0, 0, W[2,1], W[2,2], head_width=0.2, color="tab:orange", length_includes_head=true, label=L"$w_2$")
plot(cos.(θc), sin.(θc), "k--", alpha=0.8)
xlabel(L"$X_1$"); ylabel(L"$X_2$")
legend(); tight_layout()
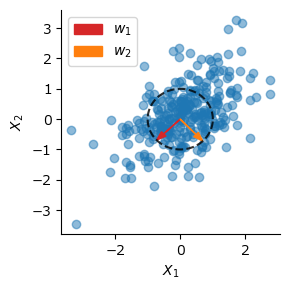
Oja則,Sanger則をまとめて一つの関数にしておこう.identity()は恒等関数である.
function HebbianPCA(X; n_components=10, η=1e-6, maxiter=200, func=identity, orthogonal=true)
# X : n x m -> Y : n_components x m
n = size(X)[1]
η /= n
X = (X .- mean(X, dims=2)) ./ std(X, dims=2) # normalization
Y = nothing
W = randn(n_components, n) # initialize weight
W ./= sqrt.(sum(W.^2, dims=2)) # normalization
for _ in 1:maxiter
Y = func.(W * X)
if orthogonal
W .+= η * (Y * X' - LowerTriangular(Y * Y') * W) # Sanger's rule
else
W .+= η * (Y * X' - Diagonal(Y * Y') * W) # Oja's rule
end
end
return Y, W
end;
非線形Hebb学習#
出力\(\mathbf{y}\)に非線形関数\(g(\cdot)\)を適用し,\(\mathbf{y}\to g(\mathbf{y})\)として置き換えることで非線形Hebb学習となる[Oja, 1997][Brito and Gerstner, 2016]. 関数HebbianPCAのfunc引数に非線形関数を渡すことで実現できる.
ToDo: 詳細
非負主成分分析によるグリッドパターンの創発#
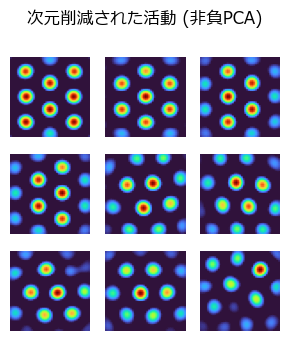
内側嗅内皮質(MEC)にあるグリッド細胞 (grid cells) は六角形格子状の発火パターンにより自己位置等を符号化するのに貢献している.この発火パターンを生み出すモデルは多数あるが,場所細胞(place cells) の発火パターンを非負主成分分析(nonnegative principal component analysis) で次元削減するとグリッド細胞のパターンが生まれるというモデルがある [Dordek et al., 2016].非線形Hebb学習を用いてこのモデルを実装しよう.なお,同様のことは非負値行列因子分解 (NMF: nonnegative matrix factorization) でも可能である.
場所細胞の発火パターン#
まず,訓練データとなる場所細胞の発火パターンを人工的に作成する.場所細胞の発火パターンはDifference of Gaussians (DoG) で近似する.DoGは大きさの異なる2つのガウス関数の差分を取った関数であり,画像に適応すればband-passフィルタとして機能する.また,DoGは網膜神経節細胞等の受容野のON中心OFF周辺型受容野のモデルとしても用いられる.受容野中央では活動が大きく,その周辺では活動が抑制される,という特性を持つ.2次元のガウス関数とDoG関数を実装する.
function gaussian2d(center, width, height, step, sigma, scale=1)
x, y = range(-width/2, width/2, length=step), range(-height/2, height/2, length=step)
f(x,y) = exp(-((x-center[1])^2 + (y-center[2])^2) / (2.0*scale*(sigma^2)))
gau = f.(x', y)
return gau ./ sum(gau)
end
function DoG(center, width=2.2, height=2.2, step=55, sigma=0.12, surround_scale=2)
g1 = gaussian2d(center, width, height, step, sigma)
g2 = gaussian2d(center, width, height, step, sigma, surround_scale)
return g1 - g2
end
DoG (generic function with 6 methods)
モデルのパラメータを設定する.
sqNp = 32 # 場所細胞の数の平方根: Np=sqNp^2
Ng = 9 # 格子細胞の数
sigma = 0.12 # 場所細胞のtuning curveの幅 [m]
surround_scale = 2 # DoGのσ²の比率
box_width = 2.2 # 箱の横幅 [m]
box_height = 2.2 # 箱の縦幅 [m]
step = 45; # 空間位置の離散化数
先にガウス関数とDoG関数がどのような見た目になるか確認しよう.
c_eg = zeros(2)
gau_eg = gaussian2d(c_eg, box_width, box_height, step, sigma)
dog_eg = DoG(c_eg, box_width, box_height, step, sigma, surround_scale);
fig, ax = subplots(2,2,figsize=(4,4),sharex="all", sharey="row")
ax[1,1].set_title("Gaussian")
ax[1,1].imshow(gau_eg, cmap="turbo", extent=(-box_width/2, box_width/2, -box_height/2, box_height/2))
ax[1,1].set_ylabel("y [m]")
ax[1,2].set_title("Difference of\n Gaussians (DoG)")
ax[1,2].imshow(dog_eg, cmap="turbo", extent=(-box_width/2, box_width/2, -box_height/2, box_height/2))
x_pos = range(-box_width/2, box_width/2, length=step)
ax[2,1].plot(x_pos, gau_eg[div(step, 2), :]/maximum(gau_eg))
ax[2,1].set_xlabel("x [m]"); ax[2,1].set_ylabel(L"$y=0$"*"の形状 (正規化)")
ax[2,2].plot(x_pos, dog_eg[div(step, 2), :]/maximum(dog_eg))
ax[2,2].set_xlabel("x [m]")
tight_layout()
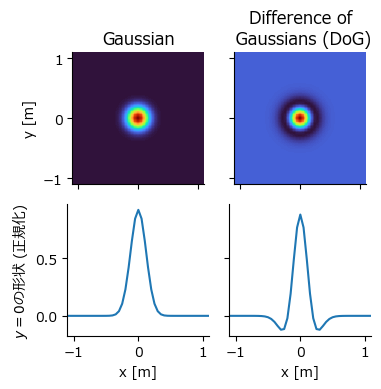
場所細胞の活動パターンを生み出す.それぞれの場所受容野の中心は空間を均等に覆うように作成する (一様分布で生み出してもよい).
x_pos = range(-box_width/2, box_width/2, length=sqNp)
y_pos = range(-box_height/2, box_height/2, length=sqNp)
centers = [[i, j] for i in x_pos for j in y_pos]
X_place = hcat([DoG(c, box_width, box_height, step, sigma, surround_scale)[:] for c in centers]...)';
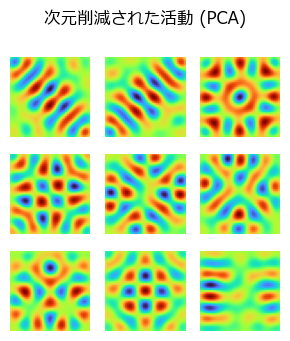
線形PCAの場合#
@time Y_pca, W_pca = HebbianPCA(X_place, n_components=Ng, η=1e-2, maxiter=5000, orthogonal=true)
Y_pca = reshape(Y_pca, (Ng, step, step));
19.396461 seconds (3.85 M allocations: 2.947 GiB, 1.45% gc time, 4.99% compilation time)
figure(figsize=(3,3.5))
suptitle("次元削減された活動 (PCA)")
for i in 1:Ng
subplot(3,3,i)
imshow(Y_pca[i, :, :], cmap="turbo")
axis("off")
end
tight_layout()
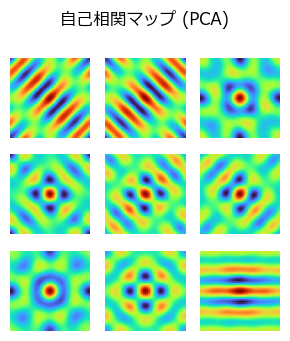
自己相関マップ(autocorrelation map)を確認する.
ToDo: 相関の計算の説明
function correlate_fft(x, y)
corr = fftshift(real(ifft(fft(x) .* conj(fft(y)))))
return corr / maximum(corr)
end;
corr_pca = [correlate_fft(Y_pca[i, :, :], Y_pca[i, :, :]) for i in 1:Ng];
figure(figsize=(3,3.5))
suptitle("自己相関マップ (PCA)")
for i in 1:Ng
subplot(3,3,i)
imshow(corr_pca[i], cmap="turbo")
axis("off")
end
tight_layout()
非負PCAの場合#
relu(x) = max(x, 0)
relu (generic function with 1 method)
@time Y_npca, W_npca = HebbianPCA(X_place; n_components=Ng, η=1e-2, maxiter=5000, func=relu, orthogonal=true);
Y_npca = reshape(Y_npca, (Ng, step, step));
19.117123 seconds (459.91 k allocations: 2.769 GiB, 1.85% gc time, 0.31% compilation time)
figure(figsize=(3,3.5))
suptitle("次元削減された活動 (非負PCA)")
for i in 1:Ng
subplot(3,3,i)
imshow(Y_npca[i, :, :], cmap="turbo")
axis("off")
end
tight_layout()
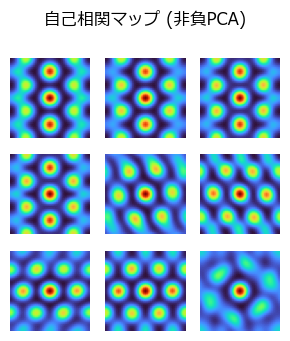
corr_npca = [correlate_fft(Y_npca[i, :, :], Y_npca[i, :, :]) for i in 1:Ng];
figure(figsize=(3,3.5))
suptitle("自己相関マップ (非負PCA)")
for i in 1:Ng
subplot(3,3,i)
imshow(corr_npca[i], cmap="turbo")
axis("off")
end
tight_layout()
ToDo: 他のgrid cellsのモデルについて
参考文献#
- BCM82
E L Bienenstock, L N Cooper, and P W Munro. Theory for the development of neuron selectivity: orientation specificity and binocular interaction in visual cortex. J. Neurosci., 2(1):32–48, January 1982.
- BL73
T V Bliss and T Lomo. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol., 232(2):331–356, July 1973.
- BG16
Carlos S N Brito and Wulfram Gerstner. Nonlinear hebbian learning as a unifying principle in receptive field formation. PLoS Comput. Biol., 12(9):e1005070, September 2016.
- CB12
Leon N Cooper and Mark F Bear. The BCM theory of synapse modification at 30: interaction of theory with experiment. Nat. Rev. Neurosci., 13(11):798–810, November 2012.
- CLO79
Leon N Cooper, Fishel Liberman, and Erkki Oja. A theory for the acquisition and loss of neuron specificity in visual cortex. Biol. Cybern., 33(1):9–28, March 1979.
- DSMD16
Yedidyah Dordek, Daniel Soudry, Ron Meir, and Dori Derdikman. Extracting grid cell characteristics from place cell inputs using non-negative principal component analysis. Elife, 5:e10094, March 2016.
- DB92
S M Dudek and M F Bear. Homosynaptic long-term depression in area CA1 of hippocampus and effects of N-methyl-D-aspartate receptor blockade. Proc. Natl. Acad. Sci. U. S. A., 89(10):4363–4367, May 1992.
- Heb49
Donald Olding Hebb. The organization of behavior: A neuropsychological theory. Wiley & Sons, 1949.
- IF20
Takuya Isomura and Karl Friston. Reverse-Engineering neural networks to characterize their cost functions. Neural Comput., 32(11):2085–2121, November 2020.
- Oja82
E Oja. A simplified neuron model as a principal component analyzer. J. Math. Biol., 15(3):267–273, 1982.
- Oja97
Erkki Oja. The nonlinear PCA learning rule in independent component analysis. Neurocomputing, 17(1):25–45, September 1997.
- Sha92
C J Shatz. The developing brain. Sci. Am., 267(3):60–67, September 1992.
- Tur08
Gina G Turrigiano. The self-tuning neuron: synaptic scaling of excitatory synapses. Cell, 135(3):422–435, October 2008.
- YHC17
Ada X Yee, Yu-Tien Hsu, and Lu Chen. A metaplasticity view of the interaction between homeostatic and hebbian plasticity. Philos. Trans. R. Soc. Lond. B Biol. Sci., March 2017.
- ZGG17
Friedemann Zenke, Wulfram Gerstner, and Surya Ganguli. The temporal paradox of hebbian learning and homeostatic plasticity. Curr. Opin. Neurobiol., 43:166–176, April 2017.